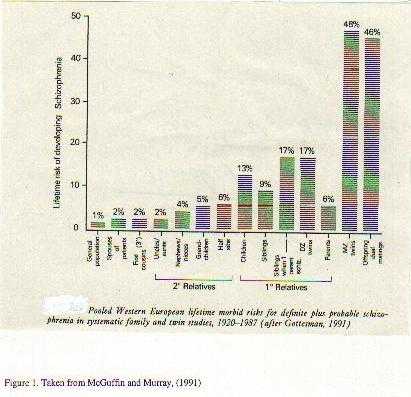
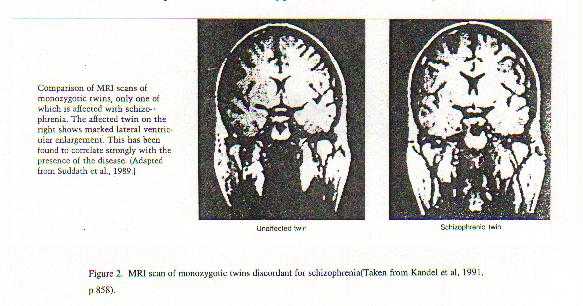
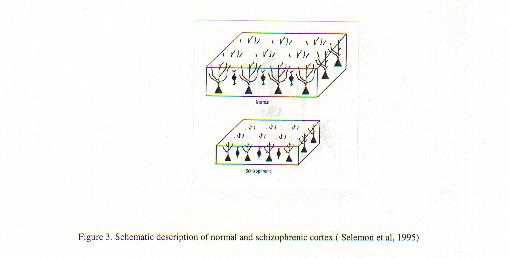
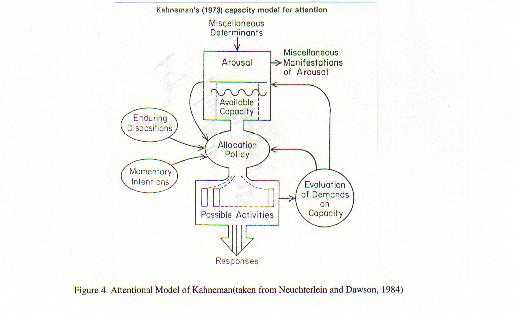
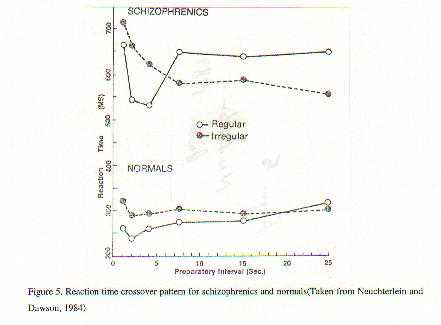
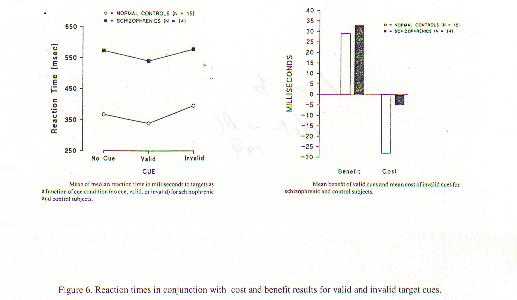
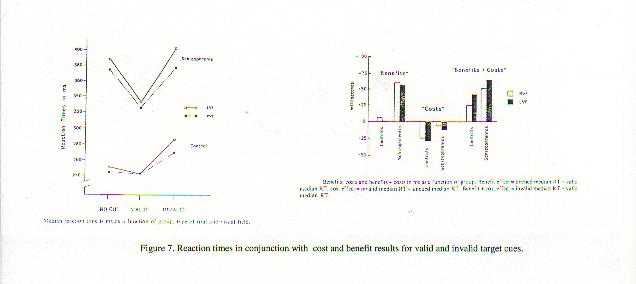
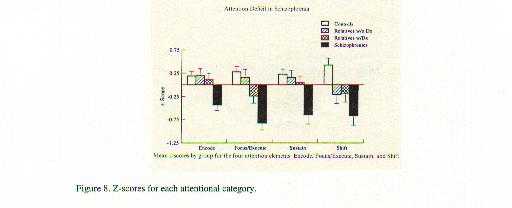
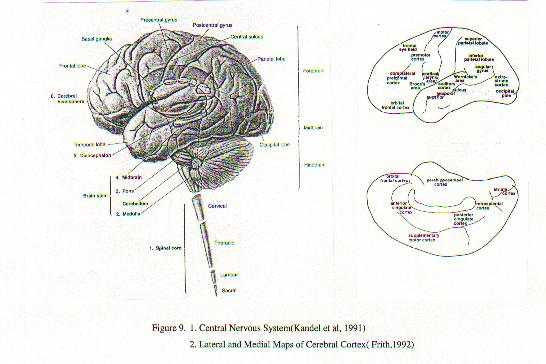
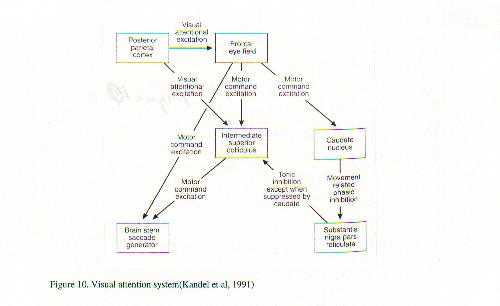
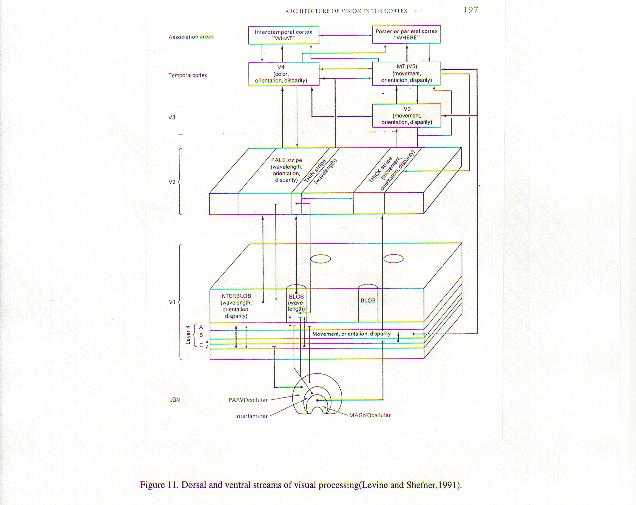
A neurobiological approach to the role of attention and automaticity was outlined by
Schneider et al(1994). This also emphasises the different processing areas in the brain
involved in effortful controlled processing and the automatic processing which have been
shown to be at the centre of the argument’s concerning the basis of schizophrenia.
They also argue that the emphasis is on anterior prefrontal cortical activation in
deliberate controlled attention relative to posterior parietal functioning when tasks
become automatised. The functional areas are shown below in figure 12.
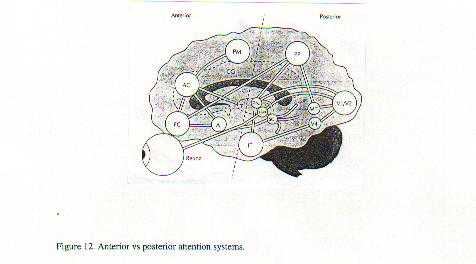
This resultant automatisation is explained as the resulting from posterior parietal
synaptic connections being strengthened or primed, as a result of task performance, in
acordance with Hebb’s synaptic strengthening rule. They note that in schizophrenia,
over time, processing continues to be maintained, predominantly, in the controlled mode.
This factor, maintenance of controlled processing, would appear to be consistent with some
of the reaction time deficits found in schizophrenics’ attentional task performance.
It has been shown that schizophrenics cannot modulate attention and they maintain
consistently high levels of arousal during selective attentional tasks(Frith et al,1988)
It has also been noted that functionality in the hippocampus increases with arousal but
when arousal reaches a critical level, hippocampal processing of information is reduced.
The earlier discussion, that had electrical stimulation of the temporal lobes causing
‘real’ experiences that did not occur in the other association areas, infers
that retrievable long term memories are stored in the temporal lobes. The role of the
hippocampus in storage of longterm memories has been demonstrated with amnesics, such as
HM, where bilateral hippocampal removal was carried out. They have lost the capacity to
encode retrievable memories for events that occurred after the damage but not for those
that occurred beforehand (Best, 1990, p189).
In terms of the priming which also results from automatic processing it is often
thought schizophrenics have excessive priming although many studies show that priming
levels are normal. It was suggested that priming might increase above normal when
neuroleptics were administered because this slowed down processing. Another interesting
possibility is the generator effect (Best, p159). When subjects are asked to generate a
word instead of just viewing it, their explicit memory in the form of recognition is
enhanced compared to priming effects. However when they do not generate words, but just
view them, priming effects are greater relative to recognition effects. If greater priming
does occur in schizophrenics as a result of a genetic characteristic it may well result in
the continuous processing of information through the non automatic ventral stream. This
process would be closer to generation than automatic processing and could be used as a
means of diminishing priming effects. This decrease in priming would create social
perceptions closer to that of normals but at the cost of the cognitive exhaustion that
would result from continuous controlled processing.
Another distinguishing feature of schizophrenia is the age of onset, typically around
the age of 20 in males and a little later in females. Is this consistent with the above
theory regarding an emphasis on controlled processing? One area of anatomical abnormality
that is often found in schizophrenics is decrease in size of the hippocampus(Bogerts et
al,1985). It would seem that, as discussed, the hippocampus is involved in controlled
processing or at least involved in encoding the information used in controlled processing,
longterm memories. It has been noted on a number of occasions that schizophrenics show
elevated arousal levels and that this causes increased excitability of hippocampal
neurons(Zahn, 1985). Myelination of the hippocampus is not completed until the late
twenties(Bencs et al, 1994) and the onset of increased retrievable longterm processing may
well be the cause of psychological problems which result in an over emphasis on controlled
processing resulting in a pathological outcome. This could be the result of vaso
constriction which seems to be associated with hippocampal activation(Sergent,1994).
Metcalfe et al, (1996) notes that the memories related to hippocampal processing are
‘cool’ memories devoid of emotional contamination but automatic processing,
which also involves the amygda, results in emotional memories or ‘hot’ memories
that can be repressed by the frontal lobes in conjunction with hippocampal memories but
this takes effort. This is a biological framework for repression. This appears to be
exactly what is happening in schizophrenia except that it is totality of automatic
processing outcome that is being suppressed rather than a specific incident.
The genetic predisposition causes high natural arousal levels which activate the
hippocampus resulting in unusual controlled processing outcomes such as greater
associative learning, as well as a less fluent interaction with the posterior parietal
system of automatic processing (see figure 13). This would create the peculiarities
observed in schizotypal personality such as difficulties in social context, magical
thinking etc. but they still maintain their psychological integrity because they do not
try to suppress this abnormality.
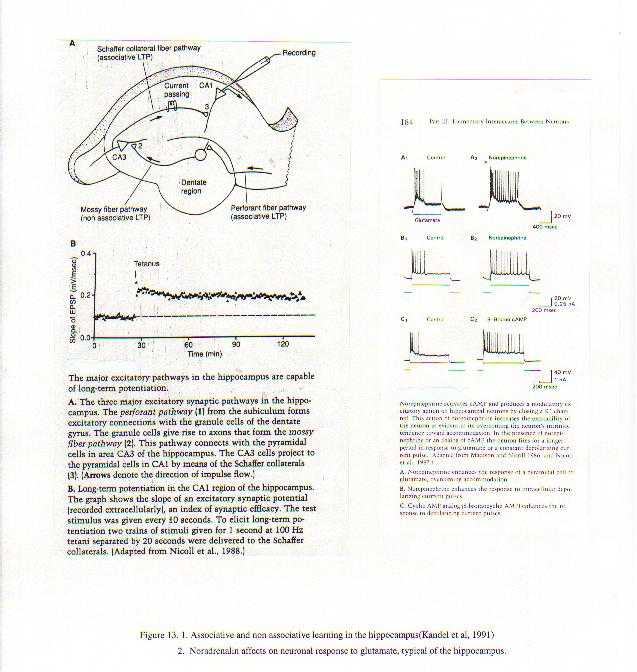
CONCLUSION
Given that thought disorder has been shown to have no genetic basis it
would seem that the onset of pathology in schizophrenia is therefore the result of faulty
strategies to cope with either developmental factors or social factors facilitated by high
arousal levels or other genetic influences resulting in increased neural excitability such
as abnormal glutamate receptor expression (Akbaran et al,1995). As arousal levels increase,
emotional salience and emotional memories, encoded into the automatic system via the
amygdala, create further psychological pressure to ‘flee’ into the controlled
mode of processing.
Attempts to ‘generate’ memories or safer realities through the organizational
capacities of the prefrontal cortex result in ‘hallucinations’ and
‘delusions’ with a detached over activation of ‘inforamtion’ encoded
in the temporal cortex ie positive symptoms. The delusions come from the higher levels of
valid spatial and temporal associations that would occur as a result of heightened
hippocampal function. It is the interpretation of these associations, especially the
erroneous attribution of causality that lead to delusions although it works both ways in
that often valid associations in regard to social interaction are denied by others ie
there have often been clinical reports where patients say even their own relatives say one
thing when they mean another. The distractibility that occurs on attentional tasks would
be generated by this focus on internal stimuli.
Negative symptoms result from a downregulation of arousal and this may result from a
learned vaso constriction capacity, which often happens after traumatic evnts, and a
passive interaction with the ventral stream of processing. This may eventually result in
cortical atrophy in the parietal areas(Zahn,1985). The lack of activation of prefrontal
areas then allows a slower more deliberate approach to task performance.
This would explain the schizophrenic dependence on concrete, object centred cognition
as opposed to the automatic, abstract and socially relevant processing that occurs in the
automatic processing mode that is associated with the dorsal posterior parietal lobe. Van
den Bosch(1994) quotes a number of patients who talk about cognitive fragmentation.
This tends to indicate that the binding capacity of the parietal cortex is playing a
secondary role to the object perception in the temporal lobe.
If the noradrenergic fibres that activated the parietal lobe neurons did so via the b -adrenergic receptors, which are widely distributed in parietal and
prefrontal cortex, we would have a situation that enabled faster performance once focus
had returned to the posterior parietal system. This would be consistent with the faster
disengagement of attention noted in schizophrenics but the slower activation of this
system results from the constant engagement of the ventral of processing. This results in
deficits in reality monitoring, central monitoring of action and social contextual
judgements, all of which rely on automatic processing.
This biological view is consistent with the attentional data presented and the
anatomical data on atrophy does not indicate irretrievable damage from cell death and
probably results from vaso constricion either self induced as a slowing mechanism or
resulting from uncontrolled excesses of noradrenalin. Acute treatment with drugs should
only be short term. A cognitive-behavioural therapy approach which explains the faulty
interactions with others as well as activities which stimulate the posterior parietal
cortex and nutritional support would seem to be the desired approach. However it may well
be that a continuum in terms of the genetic effects means that arousal level is so high
that hippocampal activity and processing will create extreme examples and this seems to be
the case with autistics.
A final note in that an interesting comparison has emerged between the schizophrenic
and the psychopath. While both have poor frontal lobe performance at certain stages, the
schizophrenic appears to be deficient in automatic processing(parietal lobe) and the
psychopath in controlled processing(temporal lobe). The schizophrenic places spiritual and
mystical interpretations on events, has a pathological lack of self awareness and a lack
of social ‘grace’. The psychopath is just the opposite and is regarded as
materialistic, selfish and socially adroit and manipulative.
REFERENCES
Akbaran, S., James, K., Potkin, S., Hagman, J., Taffazoli, A., Buney, W. and Jones,
E.(1995) Gene Expression for Glutamic Acid Decarboxylase is Reduced Without Loss of
Neurons in Prefrontal Cortex of Schizophrenics. Archives of General Psychiatry, Vol
52, 258-266.
Bencs, F., Turtle, M., Khan, Y., and Farol, P.(1994) .Myelination Of Hippocampal
Area Sharply Increases From Childhood To Adolescence To Adulthood. Archives Of General
Psychiatry, (51) 477-484.
Best,J.(1992) Cognitive Psychology 3rd Edition. West Publishing, New
York.
Bogerts, B., Meertz, E. and Schonfeldt-Bausch, R(1985). Basal Ganglia and Limbic
System Pathology in Schizophrenia: A Morphometric Study of Brain Volume and Shrinkage.
Archives of General Psychiatry, August 42(8), 784-791.
Bosch, R van den, Rombouts, R and Asma, M van(1996). What Determines Continuous
Performance Task Performance. Schizophrenia Bulletin, Vol 22, No 4.
Cai, Z. (1990). The neural mechanism of declarative memory consolidation and
retrieval: A hypothesis. Neuroscience and Biobehavioural Reviews, 14(3) 295-304.
Cannon,T, Zorrilla, L, Shtasel, D, Gur,R, Gur,R, Marco, E, Moberg,P And Price,
A.(1994). Neuropsychological Functioning In Siblings Discordant For Schizophrenia And
Healthy Volunteers. Archives Of General Psychiatry, 51:651-661
Collinge, J and Curtis, D.(1991) Decreased Hippocampal Expression of Glutamate
Receptor Gene in Schizophrenia. British Journal of Psychiatry. Vol 159, 857-859
Diagnostic and Statistical Manual of Mental Disorders(DSM IV)1994. The American
Psychiatric Association. Washington USA.
Elkins.I And Cromwell, R(1994). Priming Effects In Schizophrenia: Associative
Interference And Facilitation As A Function Of Visual Context. Journal Of Abnormal
Psychology, Vol 103, No 4,793-800.
Finkelstein, J, Cannon, T, Gur, R, Gur, R, And Moberg, P.(1997). Attentional
Dysfunctions In Neuroleptic-Naïve Neuroleptic Withdrawn Schizophrenic Patients And Their
Siblings. Journal Of Abnormal Psychology, Vol 106, No2 203-212.
Frith, C., Stylem, M., Johnstone, E. and Crow, T.(1988). Acute Schizophrenic
Patients Fail to Modulate Their Level of Attention. Journal of Psychophysiology, vol
2(3), 195-200.
Frith, C.(1992). The Cognitive Psychology of Schizophrenia. Erlbaum Press, UK.
Gold, J, Randolp, C, Carpenter, C, Goldberg,T And Weinberger, D. (1992)Forms Of
Memory Failure In Schizophrenia. Journal Of Abnormal Psychology, Vol101 No 3 487-494.
Goodale, Melvyn A. And Milner, A. David. Separate Visual Pathways For Perception And
Action. Trends In Neuroscience, Vol 15, No1, 1992.
Gorenstein, E.(1982). Frontal Lobe Function in Psychopaths. Journal of Abnormal
Psychology, Vol 91 No 5, 368-379.
Gras-Vincendon, A., Manion, J., Cramge,D., Bilik, M. and Willard-Schroeder, D., Sichel,
J. and Singer, L.(1994). Explicit Memory, Repetition Priming and Cognitive Skill
Learning in Schizophrenia. Schizophrenia Research, Vol 13, 117-126.
Gur R And Gur R.(1995) Hypofrontality In Schizophrenia:RIP. Lancet Vol 345
1383-1384.
Kandel, E., Schwartz, J. and Jessel, T.(1991). Principles of Neural Science 3rd
Edition, Appleton and Lange, NY.
Kay,S.(1990) Significance of the Positive-Negative Distinction in Schizophrenia.
Schizophrenia Bulletin, Vol 16(1), 635-652.
Levine, M. and Shefner, J.(1991)Fundamentals of Sensation and Perception 2nd
Edition. Brooks/Cole Publishing, USA.
Liotti, M, Dazzi, S And Umilta, C(1993). Deficits Of The Automatic Orienting Of
Attention In Schizophrenic Patients. Journal Of Psychiatric Research, Vol 27 No 1,
119-130.
Mesulam, M. (1990). Large Scale Neurocognitive Networks and Distributed Processing
for Attention, Language and Memory. Annals of Neurology Vol 28 No 5.
McGuffin, P and Murray, R(1991). The New Genetics of Mental Illness. Oxford:Butterworth-Heineman,
London.
Mirsky, A, Yardley,S, Jones,B, Walsh, D, And Kendler,K(1995) Analysis Of The
Attention Deficit In Schizophrenia: A Study Of Patients And Their Relatives In Ireland.
Journal Of Psychiatric Research Vol29, No1. 23-42.
Mlakar, J Jensterle J And Frith, C.(1994) Central Monitoring Deficiency And
Schizophrenic Symptoms. Psychological Medicine 24, 557-564.
Nestor, P, Faux, S, Mc Carley, R, Penhume, V, Shenton,M, And Pollak, S.(1992) Attentional
Cues In Chronic Schizophrenia: Abnormal Disengagement Of Attention. Journal Of
Abnormal Psychology, Vol 101, No4 682-689.
Posner, M And Petersen, S.(1990)The Attention System Of The Human Brain. Annual
Review Of Neuroscience, 3:25-42.
Posner, M, Early,T,Reiman,E, Pardo,P and Dwaman, M.(1988)Asymmetries in hemispheric
control of attention in schizophrenia. Archives of General Psychiatry,45, 814-821.
Raine, A and Venables, P.(1988) Enhanced P3 Evoked Potentials and Longer P3 Recovery
Times in Psychopaths.Psychophysiology, Jan 25(1) 20-38.
Raz, S.(1993) Structural Cerebral Pathology in Schizophrenia: Regional or Diffuse.
Journal of Abnormal Psychology, August Vol 102(3), 441-452.
Saykin, A J, Gur, R Gur, R, Mosely P, Mozely,L, Resnick, S Kester, B and
Stafiniak, P. (1991) Neuropsychological Function in Schizophrenia: Selective
Impairment In Memory And Learning. Archives Of General Psychiatry, Vol 48 July.
Schmand, B, Kuipers, T, van der, Gaag, M, Bosveld, J, Bulthuis, F and Jellema,M.(1994) Cognitive
Disorders and Negative Symptoms As Correlates Of Motivational Deficits In Psychotic
Patients. Psychological Medicine,24, 869-884.
Schneider W Pimm-Smith, M, Worden,M.(1994) Neurobiology of Attention and
Automaticity. Current Opinion In Neurobiology, 4:177-182.
Schroeder, Sichel,Jean Paul And Singer, Leonard.(1994) Explicit Memory, Repetition
Priming And Cognitive Skill Learning In Schizophrenia. Schizophrenia Research, Vol 13,
117-126.
Schwartz,F, Munich,R, Carr, A, Bartuch, E, Lesser, B, Rescigno, D And Viegner, B. Negative
Symptoms And Reaction Time In Schizophrenia. Journal Of Psychiatric Research, Vol 25
No 3,131-140.
Sacuzzo, D., and Schubert, D.(1981) Backward Masking as a Measure of Slow Processing
in Schizophrenia Spectrum Disorders. Journal of Abnormal Psychology Vol 90 No 4,
305-312.
Selemon, L., Rajkowska, G and Goldman-Rakic, P.(1995). Analysis of Prefrontal Area
and Occipital Area 17. Archives of General Psychiatry, October Vol 52(10), 805-818.
Sergent, J.(1994). Brain-Imaging Studies of Cognitive Functions. TINS, vol 12
No.6.
Strauss, E, Novakovic, T, Tien,A, Bylsma,F And Pearlson, G.(1991). Disengagement Of
Attention In Schizophrenia. Psychiatry Research, 37:139-146.
Taylor, S, Kornblum, S And Tandon, R(1996) Facilitation And Interference of
Selective Attention In Schizophrenia. Journal Of Psychiatric Research, Vol 30 No 4,
251-259.
Van den Bosch, R.(1994). Context And Cognition In Schizophrenia. Published In
"Advances in the Neurobiology of Schizophrenia". (Eds) Boer, J. den,
Westernberg, H. and Praag, H.. Chicester, Wiley.
Venables, P. and Patterson, T.(1978). Speech Perception and Decision Processes in
Relation to Skin Conductance and Pupillographic measures in Schizophrenia. Journal of
Psychiatric Research vol 14(1-4),183-190.
Ward, P, Catts, S, Fox, A, Michie, P, And Mcconaghy,N.(1991). Auditory Selective
Attention And Event Related Potentials In Schizophrenia. British Journal Of
Psychiatry, 158, 534-539.
Zahn T (1985) Studies of Autonomic Psychophysiology and Attention in Schizophrenia.
Schizophrenia Bulletin 14(2) 265-268.